
Introduction
Les plantes à fleurs ou angiospermes désignent le groupe des végétaux qui produisent des fleurs et des graines contenues dans un fruit pour se reproduire (contrairement aux mousses, fougères, algues, conifères, lichens,…). Elles ont colonisé de nombreux milieux, qu’ils soient terrestres ou aquatiques. Elles vivent fixées par leurs racines au sol dans un environnement auquel elles doivent être adaptées. Comment l’organisation d’une plante à fleurs lui permet-elle de vivre dans son milieu ? Nous étudierons les surfaces d’échanges avec l’air et le sol, puis la circulation de la sève, le développement, et enfin l’adaptation des plantes à leur milieu.
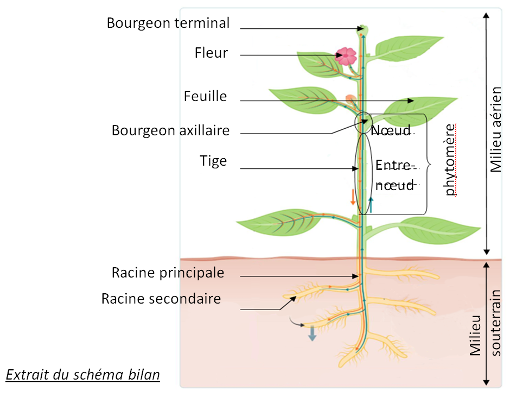
I. L’organisation d’une plante à fleurs (schéma-bilan)
Une plante à fleur est constituée :
- d’un appareil végétatif assurant la survie de l’individu. Il comprend les racines souterraines qui fixent la plante dans le sol, et la tige et les feuilles qui se développent dans le milieu aérien. Les feuilles s’insèrent au niveau des nœuds, et chaque nœud est séparé du suivant par un morceau de tige appelé entre-nœuds. Les bourgeons axillaires, qu’ils soient végétatifs pour former de nouvelles branches, ou floraux, s’insèrent au niveau des feuilles. La tige se termine par un bourgeon terminal végétatif ou floral. La tige est donc constituée d’une répétition d’une sous-unité appelée phytomère, comprenant un nœud avec les feuilles et bourgeons, et d’un entre-nœuds.
- et d’un appareil reproducteur assurant la reproduction, avec les fleurs et les graines contenues dans un fruit.
Une plante à fleurs vit donc fixée au contact de l’interface air/sol.
II. Surfaces d’échange (TP2A1)
Les plantes développent de grandes surfaces d’échange avec leur milieu extérieur :
- Les racines au niveau du sol
- Les feuilles au niveau de l’air (ou de l’eau pour les plantes aquatiques).
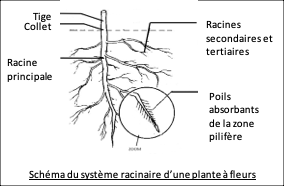
A. Souterraines
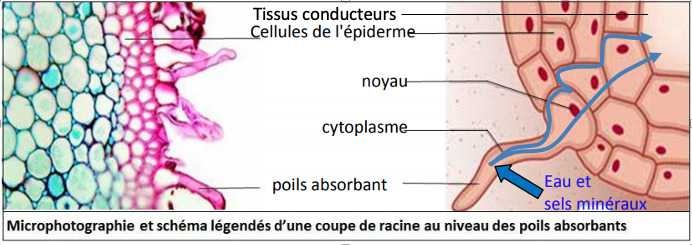
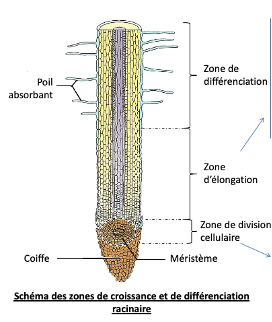
Les surfaces d’échanges souterraines se situent au niveau des racines : la racine principale se ramifie en racines secondaires et tertiaires. Près de l’extrémité de chaque racine se trouve une zone pilifère, couverte de poils absorbants très fins et nombreux (jusqu’à plusieurs milliards). Les poils absorbants sont constitués du prolongement d’une cellule de faible diamètre (Ø = 12-15µm ; longueur = qq mm) qui absorbe l’eau et les ions du sol à travers sa paroi très perméable et sa membrane plasmique. La zone pilifère se renouvelle au fur et à mesure de la croissance des racines pour maintenir une absorption élevée.
Les racines constituent donc une grande surface d’absorption d’eau et d’ions du sol grâce à leur ramification et aux poils absorbants. La surface d’absorption racinaire est ainsi estimée à plusieurs 100 m2/kg.
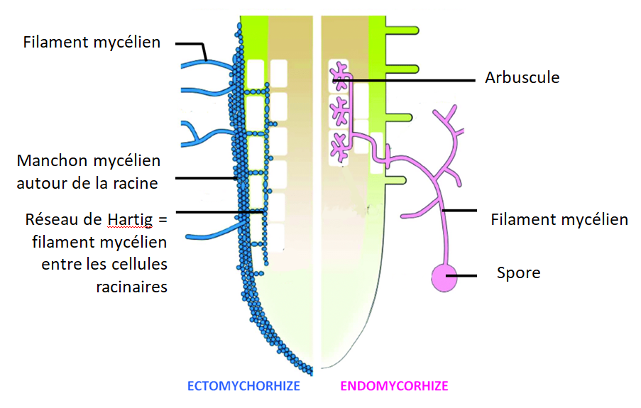
Cette surface racinaire est encore augmentée par des symbioses (= interaction intime à bénéfices réciproques) avec d’autres organismes, en particulier les filaments mycéliens de différentes espèces de champignons qui forment des mycorhizes, présentes chez ~90% des plantes à fleurs. Le champignon émet de fins filaments appelés filaments mycéliens ou hyphes. Ceux-ci s’insèrent entre les cellules racinaires hors d’une zone pilifère pour former un manchon mycélien dans le cas des ectomycorhizes(5% des plantes), ou s’insèrent entre la paroi et la membrane plasmique des cellules racinaires pour les endomycorhizes (80% des plantes terrestres). Les filaments mycéliens de ces champignons s’étendent dans le sol et augmentent la surface d’échange racinaire avec le sol. La plante reçoit l’eau et les ions captés par les filaments mycéliens, et lui fournit en retour différents nutriments qu’elle produit par photosynthèse, comme le glucose, dont le champignon, hétérotrophe (= organisme produisant sa matière organique à partir de matière minérale), a besoin pour se développer. C’est donc une symbiose.
B. Aériennes
Les surfaces d’échanges aériennes sont principalement les feuilles dont la forme est optimisée pour réaliser les échanges nécessaires à la photosynthèse :
- leur large surface plane leur permet de capter beaucoup d’énergie lumineuse nécessaire à la photosynthèse. Elles reçoivent l’énergie lumineuse principalement au niveau de leur face supérieure où se situent les hautes cellules chlorophylliennes du parenchyme palissadique qui contiennent de nombreux chloroplastes. La feuille est recouverte sur chaque face d’un épiderme (couche d’une seule épaisseur de cellules en surface) recouvert d’une cuticule (couche externe cireuse imperméable) qui protège les pigments photosynthétiques contre les UV qui les dégradent. Cette cuticule est imperméable, ce qui empêche les échanges gazeux, sauf au niveau des stomates.
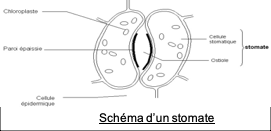
- les transferts de gaz participant à la photosynthèse (CO2, O2, H2O) sont réalisés au niveau des stomates : ce sont des structures situées dans l’épiderme, constituées d’une ouverture (ostiole) encadré par 2 cellules stomatiques réniformes (en forme de rein) = 2 cellules de garde ; un stomate se trouve au contact d’une chambre sous-stomatique, un espace rempli d’air entouré de cellules chlorophylliennes dans le parenchyme lacuneux. Les stomates permettent les échanges gazeux : quand ils sont ouverts, le CO2nécessaire à la photosynthèse entre, et le O2 et l’eau libérés au cours de la photosynthèse sortent entre la chambre sous-stomatique et l’air extérieur par diffusion au niveau des stomates. Les stomates sont présents souvent majoritairement sur la face inférieure des feuilles, ce qui permet de limiter les pertes d’eau, la face inférieure étant moins chaude et donc moins soumise au dessèchement.
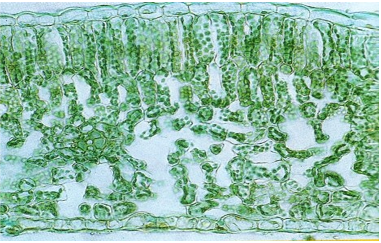
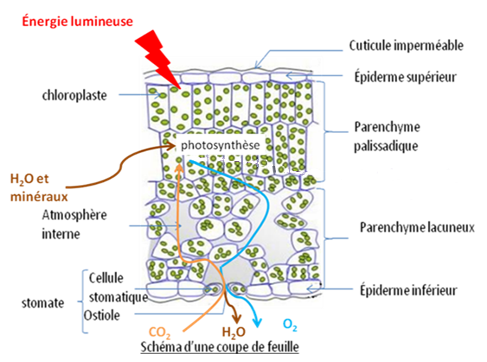
Photographie d’une coupe transversale de feuille et son interprétation schématique (x100)
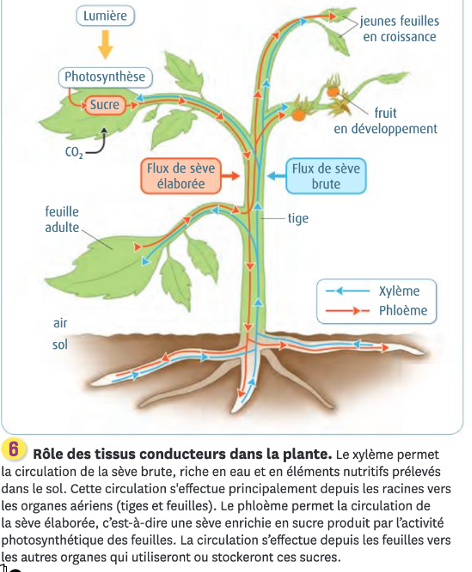
II. Circulations (TP2A2)
L’eau et les minéraux sont prélevés par les racines dans le sol, alors que la photosynthèse a lieu au niveau des feuilles dans l’air. L’eau et les minéraux nécessaires à la photosynthèse doivent donc être amenés des racines aux feuilles, et la matière organique synthétisée dans les feuilles doit être amenée jusqu’aux racines pour leur croissance et la constitution de réserves. Les circulations de matière entre l’appareil végétatif souterrain et aérien s’effectuent grâce à un double système de conduction de sève par deux types de tissus conducteurs différents.
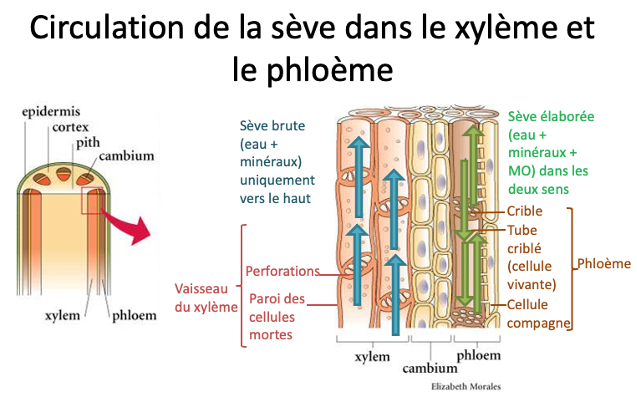
A. Le xylème et la sève brute
L’eau et les minéraux sont prélevés par les racines dans le sol. Elle forme dans la racine la sève brute. Elle circule latéralement dans la racine jusqu’au xylème. Le xylème est un tissu conducteur constitué d’un ensemble de cellules conductrices mortes dont il ne reste que la paroi imprégnée de lignine, une molécule qui rigidifie la paroi, déposée en anneaux ou en spirales. Cela forme des vaisseaux dans lesquels la sève brute circule vers le haut des racines en passant par les tiges jusqu’aux feuilles, aux bourgeons, aux fleurs et fruits.
B. Le phloème et la sève élaborée
Les cellules chlorophylliennes utilisent l’eau et les minéraux pour la photosynthèse. Grâce à cela, elles produisent de nombreuses molécules (glucides, acides aminés des. Avec de l’eau, elles forment la sève élaborée qui circule dans le phloème : c’est un tissu conducteur composé essentiellement de tubes criblés, des cellules vivantes dont la paroi est imprégnée de cellulose. La sève élaborée peut circuler vers le haut ou vers le bas selon les besoins de la plante, des organes sources que sont les parties chlorophylliennes, vers les organes puits en général non chlorophylliens (racines, fleurs écloses, organes de stockage de réserves : fruit, tubercule = racine gorgée de réserve, bulbe, oignon…).
IV. Développement (TP2A3)
A. Croissance et différenciation
Le développement d’une plante associe :
La croissance qui permet l’allongement de la tige et des racines. Elle se fait de 2 façons :
- Par la multiplication cellulaire : des cellules totipotentes (= cellules indifférenciées capables de se diviser et de se différencier en n’importe quel type de cellule) se divisent activement par mitoses dans des zones spécialisées dans la croissance et le développement, situées à l’extrémité des racines ou des bourgeons terminaux et axillaires, les méristèmes caulinaires (= de la tige) et racinaires. Rq : des méristèmes secondaires (cambium) permettent l’accroissement en largeur en produisant des tissus conducteurs année après année.
- Par l’élongation cellulaire : après division, les cellules s’allongent dans la zone d’élongation racinaire, ou dans l’entre-nœud de la tige.
et la différenciation d’organes (tiges, feuilles, fleurs, racines) à partir du méristème : c’est l’organogenèse (= formation des organes). Elle s’appuie sur la différenciation des cellules végétales après leur division, qui leur permet d’acquérir une structure en lien avec leur fonction. Les cellules du méristème apical forment par exemple des ébauches de feuilles. Rq : les cellules racinaires peuvent se dédifférencier pour former des méristèmes à l’origine des racines secondaires.
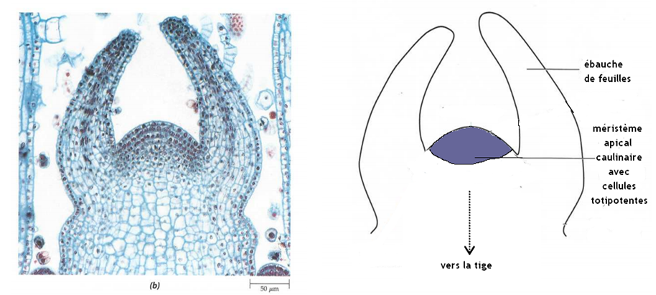
Coupe longitudinale de bourgeon caulinaire
Ce développement conduit à une organisation modulaire en phytomères qui sont des structures répétitives constituées de tige + feuille (+ bourgeon axillaire).
B. Contrôle
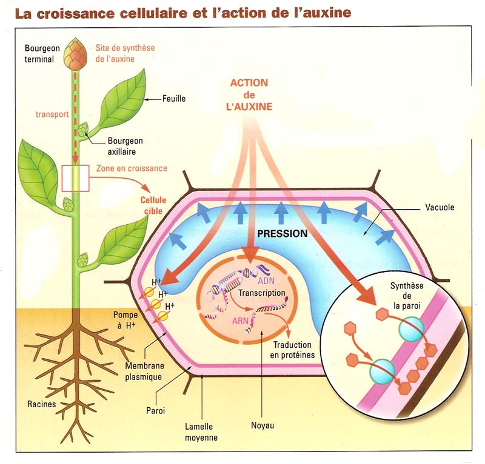
Ce développement est influencé par les conditions du milieu. Il est contrôlé par des hormones végétales = phytohormones (= molécules végétales produite par des cellules végétales et qui agit sur d’autres cellules végétales), dont les concentrations dépendent des conditions de l’environnement. Ainsi, l’auxine (= AIA = acide indole acétique) est une phytohormone produite par le bourgeon terminal qui stimule l’élongation des cellules à des concentrations moyennes.
Ainsi, lors d’un éclairement latéral anisotrope (=non homogène), la tige s’oriente vers la lumière : c’est le phototropisme. L’auxine migre vers la zone la moins éclairée de la tige, ce qui favorise l’élongation de la tige du côté opposé à la lumière, et sa courbure vers la lumière. De même, les racines s’orientent vers le bas grâce à la gravité : c’est le gravitropisme. Ici, l’auxine migre vers les cellules du bas de la racine, et atteint des concentrations suffisantes pour inhiber l’élongation des cellules, ce qui favorise la courbure de la racine vers le bas. Le développement peut aussi être contrôlé par d’autres facteurs de l’environnement, comme le vent, la quantité de lumière disponible, … Tout ceci permet à la plante de s’adapter à son environnement.
D’autres phytohormones existent, comme les cytokinines qui développent les bourgeons axillaires. L’éthylène est produit pour faire mûrir les fruits, lors de la chute des feuilles en automne, ou dans toute situation de stress.
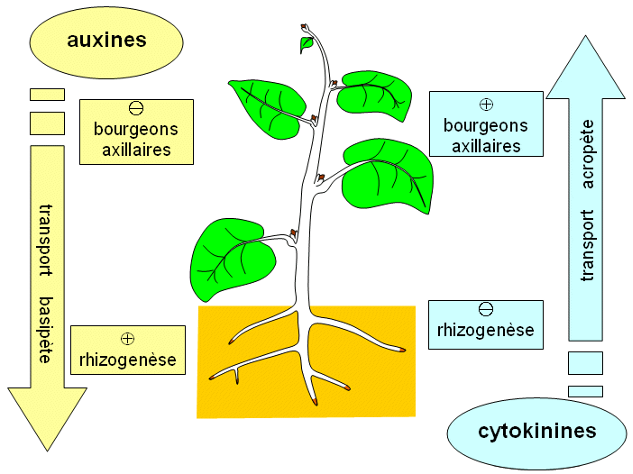
Effets de l’auxine et des cytokinines sur la croissance végétale
V. Adaptations à l’environnement (TP2A4)
Les plantes sont fixées au sol et sont adaptées à des environnements variables, principalement par leur surface d’échange, de façon transitoire par leur physiologie, ou de façon permanente par leur morphologie.
A. La sécheresse
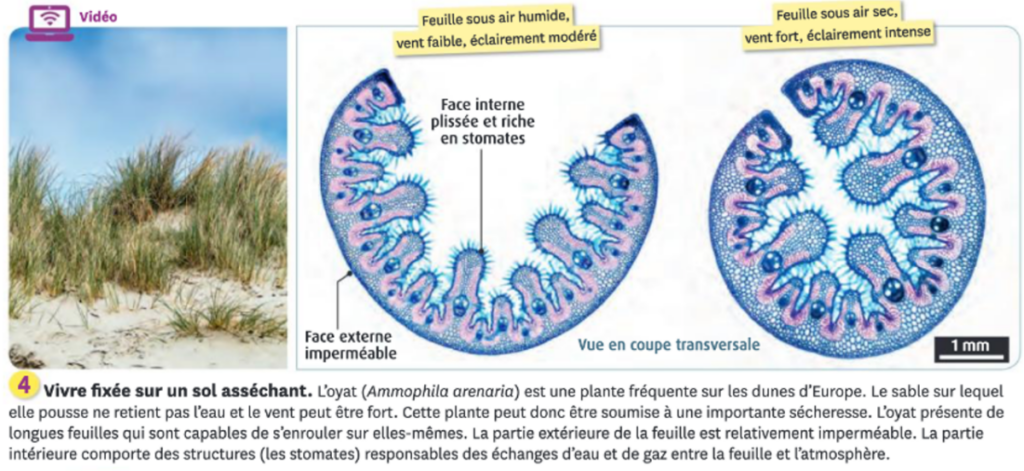
La sécheresse peut être due à des faibles précipitations, à la chaleur et/ou au vent. Chez la plupart des plantes, les stomates se ferment pendant un épisode sec, ce qui empêche les pertes d’eau par évapotranspiration (libération d’eau au niveau des stomates lors de la photosynthèse) mais empêche aussi la photosynthèse. Pour permettre une photosynthèse même en milieu sec, certaines plantes ont développé des adaptations à la sécheresse qui nécessite de limiter les pertes en eau :
- Limiter les surfaces d’échange des feuilles avec l’atmosphère sèche, notamment grâce à :
- des stomates situés sur la face inférieure (ex : laurier-rose), parfois dans des cryptes pilifères (= munies de poils) qui captent l’humidité
- une surface en contact avec l’air réduite (ex : feuilles enroulées de l’Oyat) ;
- et une face supérieure couverte d’une cuticule imperméable épaisse
- Augmenter l’absorption d’eau en augmentant la surface d’échange des racines qui se développent davantage grâce à l’auxine.
B. Le froid
Le froid et en particulier le gel en hiver fait éclater les cellules. Pour réduire leur surface d’échange et limiter les pertes thermiques, les plantes en milieu alpin ont un port réduit de plus petite taille, et des petites feuilles. Les Conifères ont des aiguilles pour limiter les échanges thermiques. Plus généralement, les plantes des milieux tempérés ont développé des adaptations pour éviter que les feuilles ne gèlent en hiver (évitement). Les arbres à feuilles caduques perdent leurs feuilles en hiver grâce à une phytohormone, l’éthylène, qui favorise la sénescence et la chute des feuilles à l’automne. Les bourgeons protégés par des écailles (feuilles épaisses et imperméables) reformeront les feuilles et les fleurs au printemps suivant. Certaines plantes passent l’hiver dans le sol qui protège du froid et du gel :
- Certaines plantes vivaces (qui survivent plusieurs années) passent l’hiver sous forme d’organes souterrains (dans le sol, la plante est protégée du froid). Ces organes souterrains sont variés : rhizome, bulbes, tubercules,… Au printemps suivant, les réserves seront utilisées pour reformer la plante.
- Les plantes annuelles (qui ne vivent qu’une année) passent la mauvaise saison sous forme de graines qui contiennent peu d’eau et résistent au froid. La plantule miniature à l’intérieure de la graine se développera au printemps suivant.
Les plantes produisent aussi de façon temporaire des molécules qui leur permettent de mieux résister aux baisses de températures en hiver.
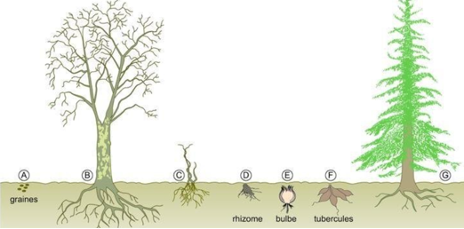
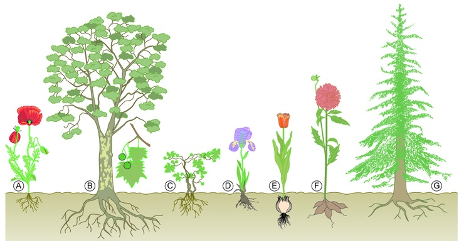
Plantes vivaces et annuelles en hiver (en haut) et en été (en bas)
Conclusion
Les plantes à fleurs sont adaptées à la vie fixée : elles possèdent de larges surfaces d’échanges (feuilles, racines) qui leur permettent de réaliser la photosynthèse. Les ions et molécules absorbés et synthétisés sont transportés dans la plante sous forme de sève par 2 types de tissus conducteurs : le xylème (sève brute) et le phloème (sève élaborée). Elles se développent sous l’effet des conditions du milieu qui déterminent la répartition des phytohormones, leur permettant de s’adapter à différents environnements.
Pour aller plus loin: