A. Le code génétique
La séquence des nucléotides de l’ARNm gouverne la séquence des acides aminés dans la protéine selon un système de correspondance : le CODE GÉNÉTIQUE. Un triplet (ou codon) de nucléotides (= groupe de 3 nucléotides) code un acide aminé (sauf les codons-stop : voir IIIB.). Le code génétique possède 3 propriétés, il est :
- UNIVOQUE (non ambigu) : à chaque codon de nucléotides correspond un seul acide aminé. (ex: UCU code uniquement la sérine)
- REDONDANT = DÉGÉNÉRÉ : comme 64 codons de nucléotides codent 20 acides aminés, plusieurs codons correspondent à un même acide aminé. (ex: UCU, UCC, UCA et UCG codent tous les 4 la sérine)
- UNIVERSEL : c’est le même pour presque tous les êtres vivants.

Une animation pour apprendre à utiliser le code génétique: http://svt.pages.ac-besancon.fr/codegenetique/. Cliquer sur les nucléotides d’un codon successivement et lire le résultat.
B. La synthèse d’une protéine à partir de l’ARNm
La synthèse d’une protéine à partir de l’ARNm est la TRADUCTION. Elle est réalisée par les RIBOSOMES, des petits éléments granuleux présents dans le cytoplasme; les ribosomes se lient à l’ARNm au niveau du codon d’initiationAUG(initiation) puis progressent le long de l’ARNm en associant un acide aminé à chaque codon selon le code génétique et en le liant au reste de la chaine d’acides aminés(élongation). Au niveau du codon-stop, aucun acide aminé n’est ajouté, et la chaine d’acides aminés est libérée dans le cytoplasme, et le ribosome se détache de l’ARNm, ce qui termine la traduction(terminaison).La chaine d’acides aminés se replie ensuite pour former une protéine dans le cytoplasme.
Plusieurs ribosomes traduisent en même temps un même ARNm (polysome) : c’est l’amplification.

Un brin d’ARNm traduit par plusieurs ribosomes en même temps (MET)

Schéma des étapes de la traduction
Une vidéo sur les étapes de la traduction. Tout ce qui n’a pas été abordé en cours est hors programme (ARN de transfert des ribosomes, sous-unités, sites A et P de la grosse sous-unité).








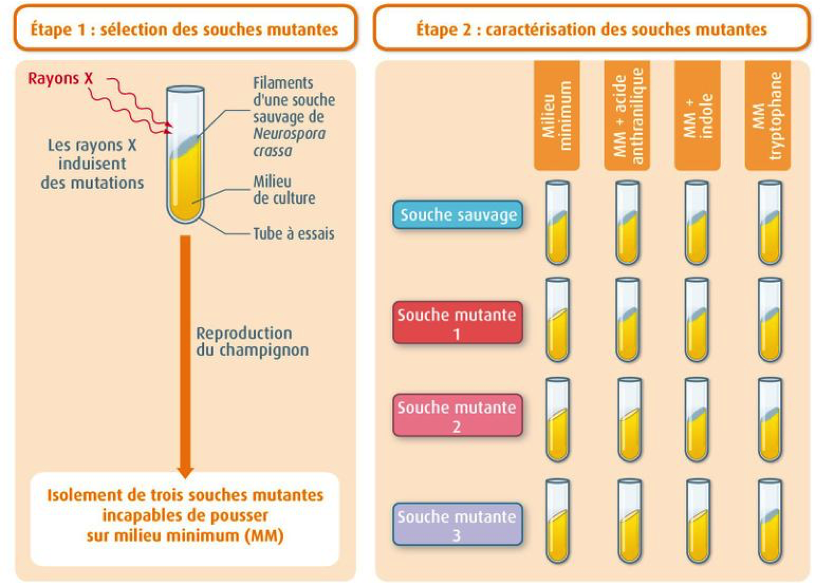
 Expérience historique de Beadle et Tatum: La mutation d’un gène entraine la modification d’une protéine (une enzyme) qui entraine la modification d’un caractère héréditaire (la capacité à synthétiser une molécule et à pousser sur un milieu supplémenté ou non en cette molécule)
Expérience historique de Beadle et Tatum: La mutation d’un gène entraine la modification d’une protéine (une enzyme) qui entraine la modification d’un caractère héréditaire (la capacité à synthétiser une molécule et à pousser sur un milieu supplémenté ou non en cette molécule)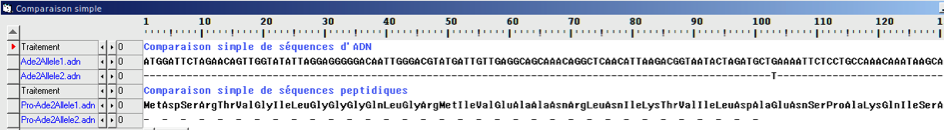






.png)